

jueves, 3 de julio de 2008
jueves, 26 de junio de 2008
Por Victor Cárdenas y Matías Santana
La membrana plasmática o citoplasmática es una estructura laminar que engloba a las células, define sus límites y contribuye a mantener el equilibrio entre el interior y el exterior de éstas. Además, se asemeja a las membranas que delimitan los orgánulos de células eucariotas.

Esta compuesta por una que sirve de "contenedor" para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica. Está formada principalmente por lípidos y proteínas. La principal característica de esta barrera es su permeabilidad selectiva, lo cual le permite "seleccionar" las moléculas que entran y salen de la célula. Tiene un grosor aproximado de 75 Å. No es visible al microscopio óptico pero sí al microscopio electrónico, donde se pueden observar dos capas oscuras laterales y una central más clara.
En las células procariotas y en las de eucariontes osmótrofos como plantas y hongos, se sitúa bajo otra capa, denominada pared celular.
Composición química
La composición química de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentren, pero se puede estudiar de forma general. La membrana plasmática está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y glúcidos unidos covalentemente a lípidos o proteínas. Las moléculas más numerosas son las de lípidos, ya que se cree que por cada 50 lípidos hay una proteína. Sin embargo, la proteína, debido a su mayor tamaño, representan aproximadamente el 50% de la masa de la membrana.
Proteínas. El porcentaje de proteínas oscila entre un 20% en la vaina de mielina de las neuronas y un 70% en la membrana interna mitocondrial; el 80% son intrínsecas, mientras que el 20% restantes son extrínsecas. Las proteínas son responsables de las funciones dinámicas de la membrana, por lo que cada membrana tienen una dotación muy específica de proteínas; las membranas intracelulares tienen una elevada proporción de proteínas debido al elevado número de actividades enzimáticas que albergan. Hay proteínas con diferentes funciones en la membrana plasmática: transportadoras, conectoras (conectan la membrana con la matriz extracelular o con el interior), receptoras (encargadas del reconocimiento celular y adhesión) y enzimas.
Lípidos. El 98% son anfipáticos, es decir que presentan un extremo hidrófilo (que tiene afinidad e interacciona con el agua) y un extremo hidrofóbico (que repele el agua). Los más abundantes son los fosfoglicéridos (fosfolípidos) y los esfingolípidos, que se encuentran en todas las células; le siguen los glucolípidos, así como esteroides, como el colesterol. Estos últimos no existen o son escasos en las membranas plasmáticas de las células procariotas. Existen también grasas neutras, que son lípidos no anfipáticos pero sólo representan un 2% del total de lípidos de membrana.
Fosfoglicéridos. Tienen una molécula de glicerol con la que se esterifica un ácido fosfórico y dos ácidos grasos de cadena larga; los principales fosfoglicéridos de membrana son la fosfatidiletanolamina o cefalina y la fosfatidilcolina o lecitina.
Esfingolípidos. Son lípidos de membrana constituidos por ceramida (esfingosina + ácido graso); solo la familia de la esfingomielina posee fósforo; el resto poseen glúcidos y se denominan por ello glucoesfingolípidos o, simplemente glucolípidos. Los cerebrósidos poseen principalmente glucosa, galactosa y sus derivados (como N-acetilglucosamina y N-acetilgalactosamina). Los gangliósidos contienen una o más unidades de ácido N+acetilneuramínico (ácido siálico).
Colesterol. El coleterol representa un 23% de los lípidos de membrana. Sus moléculas son pequeñas y más anfipáticas en comparación con otros lípidos. Se dispone con el grupo hidroxilo hacia el exterior de la célula (ya que ese hidroxilo interactúa con el agua). El colesterol es un factor importante en la fluidez y permeabilidad de la membrana ya que ocupa los huecos de la membrana. A mayor cantidad de colesterol, menos permeable y fluida es la membrana. Se ha postulado que los lípidos de membrana se podrían encontrar en dos formas: como un líquido bidimensional, y de una forma más estructurada, en particular cuando están unidos a algunas proteínas formando las llamadas balsas lipídicas. Se cree que el colesterol podría tener un papel importante en la organización de estas últimas.
Glúcidos: están en la membrana unidos covalentemente a proteínas o a lípidos. Pueden ser polisacáridos u oligosacáridos. Se encuentran en el exterior de la membrana formando el glicocalix. Representan el 8% del peso seco de la membrana plasmática.
Estructura
Artículo principal: Aspectos estructurales de la membrana plasmática
Antiguamente se creía que la membrana plásmatica era un conjunto estático formado por las siguientes capas: Proteínas/Lípidos/Lípidos/Proteínas. Hoy en día se concibe como una estructura dinámica.
Esquema de una membrana celular. Según el modelo del mosaico fluido, las proteínas (en rojo y naranja) serían como "icebergs" que navegarían en un mar de lípidos (en azul). Nótese además que las cadenas de oligosacáridos (en verde) se hallan siempre en la cara externa, pero no en la interna.
Su modelo estructural es conocido como mosaico fluido. El "mosaico fluido" es un término acuñado por S. J. Singer y G. L. Nicholson en 1972. Este consiste en una bicapa lipídica complementada con diversos tipos de proteínas. La estructura básica se mantiene unida mediante uniones no covalentes.
Las proteínas de la membrana plasmática se pueden clasificar según cómo se dispongan en la bicapa lipídica
Proteínas integrales: Embebidas en la bicapa lipídica, atraviesan la membrana una o varias veces, asomando por una o las dos caras (proteínas transmembrana); o bien mediante enlaces covalentes con un lípido o a un glúcido de la membrana. Su aislamiento requiere la ruptura de la bicapa.
Proteínas periféricas: A un lado u otro de la bicapa lipídica, pueden estar unidas débilmente por enlaces no covalentes. Fácilmente separables de la bicapa, sin provocar su ruptura.
Los glúcidos se hallan asociados mediante enlaces covalentes a lípidos, proteínas y generalmente forman parte de la matriz extracelular.
Otras sustancias pueden estar asociadas a esta estructura básica como diversos tipos de glúcidos que pueden unirse de forma covalente a lípidos (glucolípidos) o a proteínas (glucoproteínas). Las cadenas de estos glúcidos se disponen hacia el medio extracelular por la cara externa de la membrana y constituyen el glucocálix o matriz extracelular.
Esta estructura general -modelo unitario- se presenta también en las membranas de diversos orgánulos del interior de la célula: los del sistema de endomembranas, tales como retículo endoplasmático, aparato de Golgi y envoltura nuclear, y los de otros orgánulos, como las mitocondrias y los plastos, que proceden de endosimbiosis.
Funciones
La función básica de la membrana plasmática reside en mantener el medio intracelular diferenciado del entorno. Esto es posible gracias a la naturaleza aislante en medio acuoso de la bicapa lipídica y a las funciones de transporte que desempeñan las proteínas. La combinación de transporte activo y transporte pasivo hacen de la membrana plasmática una barrera selectiva que permite a la célula diferenciarse del medio.
Los esteroides, como el colesterol, tienen un importante papel en la regulación de las propiedades físico-químicas de la membrana regulando su resistencia y fluidez.
En el componente proteico reside la mayor parte de la funcionalidad de la membrana, las proteínas realizan funciones específicas y podemos clasificarlas según su función en:
Estructurales: estas proteínas hacen de "eslabón clave" uniéndose al citoesqueleto y la matriz extracelular.
Receptores de membrana: que se encargan de la recepción y transducción de señales químicas.
Transportadoras a través de membrana: mantienen un gradiente electroquímico mediante el transporte de membrana de diversos iones.
Estas a su vez pueden ser:
Proteínas transportadoras: Son enzimas con centros de reacción que sufren cambios conformacionales.
Proteínas de canal: Dejan un canal hidrofílico por donde pasan los iones.
En el transporte transmembrana podemos hablar de:
Transporte pasivo: Se produce sin consumo de energía y a favor de gradiente electroquímico, o de concentración.
Transporte activo: Se produce con consumo de energía y en contra de gradiente electroquímico.
El componente glucídico forma el glucocáliz, con funciones de cierta protección ante agresiones mecánicas y químicas, y la que parece más importante ya que permite diferenciar el exterior celular permitiendo un reconocimiento intercelular.
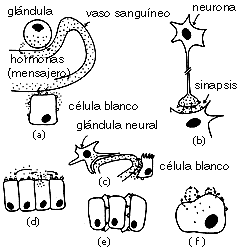
En citología, el término receptores designa a las proteínas que permiten la interacción de determinadas sustancias con los mecanismos del metabolismo celular. Los receptores son proteínas o glicoproteínas presentes en la membrana plasmática, en la membrana de las organelas o en el citosol celular, a las que se unen específicamente otras sustancias químicas llamadas moléculas señalizadoras, como las hormonas y los neurotransmisores.
La unión de una molécula señalizadora a sus receptores específicos desencadena una serie de reacciones en el interior de las células (Transducción de señal), cuyo resultado final depende no solo del estímulo recibido, sino de muchos otros factores, como el estadio celular, la situación metabólica de la célula, la presencia de patógenos, el estado metabólico de la célula, etc.
Reconocimiento de la hormona por los receptores transmembrana El reconocimiento de la estructura química de una hormona por el receptor de la hormona utiliza los mismos mecanismos de enlace no covalente como los puentes de hidrógeno, fuerzas electrostáticas, fuerzas hidrófobas y de Van der Waals. La equivalencia entre la unión hormona-receptor y la hormona libre es igual a: [H] + [R] <-> [HR], con
[R]=receptor; [H]=hormona libre; [HR]=receptor unido a la hormona
Lo importante de la fuerza de la señal transmitida por el receptor es la concentración de complejos hormona-receptor, que es definida por la afinidad que existe entre la hormona con su receptor, por la concentración de la hormona y por la concentración del receptor. La concentración de hormona circulante es el punto principal de la fuerza de la señal, siempre que los otros dos valores sean constantes. En reacciones rápidas, la producción de hormonas por las células puede almacenarse en forma de prohormonas, y rápidamente transformarse y liberarse cuando sea necesario.
También la célula puede modificar la sensibilidad del receptor, por ejemplo por la fosforilación. También por la variación del número de receptores que pueden modificar la fuerza total de señalización en el interior de la celula.
Receptores nucleares
Los receptores nucleares o citoplasmáticos son proteínas solubles localizadas en el citoplasma o en el núcleo celular. La hormona que pasa a través de la membrana plasmática, normalmente por difusión pasiva, alcanza el receptor e inicia la cascada de señales. Los receptores nucleares son activadores de la transcripción activados por ligandos, que se transportan con el ligando u hormona, que pasan a través de la membrana nuclear al interior del núcleo celular y activan la transcripción de ciertos genes y por lo tanto la producción de una proteína.
Los ligandos típicos de los receptores nucleares son hormonas lipofílicas como las hormonas esteroideas, por ejemplo la testosterona, la progesterona y el cortisol, derivados de la vitamina A y vitamina D. Estas hormonas desempeñan una función muy importante en la regulación del metabolismo, en las funciones de muchos órganos, en el proceso de desarrollo y crecimiento de los organismos y en la diferenciación celular. La importancia de la fuerza de la señal es la concentración de hormona, que está regulada por:
Biosíntesis y secreción de hormonas por los órganos endocrinos: Por ejemplo el hipotálamo recibe información, tanto eléctrica como bioquímica. El hipotálamo produce factores liberadores de hormonas que actúan sobre la hipófisis y activa la producción de hormonas hipofisarias, las cuales activan los órganos endocrinos que finalmente producen las hormonas para los tejidos diana. Este sistema jerarquizado permite la amplificación de la señal original que procede del hipotálamo. La liberación de hormonas enlentece la producción de estas hormonas por medio de una inhibición reactiva (feedback), para evitar una producción aumentada.
Disponibilidad de la hormona en el citoplasma: Muchas hormonas pueden ser convertidas en formas de depósito por la célula diana para su posterior uso. Este reduce la cantidad de hormona disponible.
Modificación de las hormonas en el tejido diana: Algunas hormonas pueden ser modificadas por la célula diana, de modo que no activan el receptor hormonal y así reducen la cantidad de hormonas disponibles.
Los receptores nucleares que son activados por hormonas activan receptores específicos del ADN llamados elementos sensibles a hormonas (HREs, del inglés Hormone Responsive Elements), que son secuencias de ADN que están situados en la región promotora de los genes que son activados por el complejo hormona receptor. Como este complejo activa la transcripción de determinados genes, estas hormonas también se llaman inductores de la expresión genética. La activación de la transcripción de genes es mucho más lenta que las señales que directamente afectan a proteínas ya existentes. Como consecuencia, los efectos de hormonas que se unen a receptores nucleares se producen a largo plazo. Sin embargo la señal de transducción a través de receptores solubles afecta sólo a algunas proteínas. Los detalles de la regulación genética todavía no son del todo conocidos. Todos los receptores nucleares tienen una estructura modular similar:
Receptores esteroideos
Los receptores esteroideos son un subtipo de receptores nucleares localizados permanentemente en el citoplasma. En ausencia de hormona esteroidea, los receptores están unidos en un complejo denominado complejo aporreceptor, que contiene proteínas chaperonas o carabina, también conocidas como proteínas de choque térmico o de calor (HSPs del inglés Heat Shock Proteins). Las HSPs son necesarias en la activación del receptor porque ayuda a cambiar su conformación que le permite unirse a la secuencia de bases del ADN.
Los receptores esteroides también pueden tener un efecto represivo sobre la expresión genética cuando el dominio de transactivación esté escondido, por lo que no se puede activar la transcripción. Además la actividad del receptor esteroideo puede ser aumentada por la fosforilación de residuos de serina en su N-terminal, como resultado de otras formas de señal de transducción, por ejemplo como por un factor de crecimiento. Este comportamiento es llamado crosstalk.
RXS y receptores huérfanos
Estos receptores moleculares pueden ser activados por:
Una hormona clásica que entra en la célula por difusión.
Una hormona que fue sintetizada en la célula, como por ejemplo retinol, de un precursor o prohormona, que puede ser transportada hacia la célula a través del torrente sanguíneo.
Una hormona que fue completamente sintetizada en el interior de la célula por ejemplo, las prostaglandinas.
Estos receptores están localizados en el núcleo y no están acompañados de proteínas carabina. En ausencia de hormona, se une a su secuencia específica de ADN inactivando un gen. Cuando se activan por las hormonas, se activa la transcripción de genes que estaban reprimidos.

Calcio como segundo mensajero
El calcio actúa como una molécula de señal dentro de la célula. Cuando el calcio es liberado y por lo tanto es activo, actúa en un espacio muy limitado de tiempo. Por lo tanto la concentración de ion calcio dentro de la célula es muy bajo normalmente. El calcio está almacenado dentro de orgánulos, normalmente en el retículo endoplásmico o retículo sarcoplásmico en las células musculares, donde está rodeado de moléculas parecidas a la calreticulina.
El receptor InsP3 puede transportar calcio a través de la interacción con inositoltrifosfato en la cara citoplasmática. Está formado por cuatro subunidades idénticas.
El receptor rianodin, llamado así por el vegetal alcaloide rianodin, es similar al receptor del insP3 y estimula el transporte del calcio al interior del citoplasma por el reconocimiento del calcio en lugares citosólicos, de este modo se establece un mecanismo de retroalimentación, en el que una pequeña cantidad de calcio en el citosol cerca del receptor, puede provocar la liberación de más calcio. Esto es especialmente importante en neuronas y células musculares. En las células del corazón y del páncreas otro segundo mensajero, el ADP cíclico de ribosa, forma parte de la activación del receptor.
La localización y el tiempo limitado del calcio en el citoplasma se llama ola de calcio. La formación de la oleada es debida a:
El mecanismo de retroalimentación positivo (feedback) del receptor rianodin.
La activación de fosfolipasa por el calcio, el cual estimula la producción de inositol trifosfato que vuelve a activar al receptor InsP3.
Función del calcio
El calcio está implicado en múltiples procesos como la contracción muscular, la liberación de neurotransmisores desde las terminaciones nerviosas, la visión en las células de la retina, proliferación, secreción, funcionamiento del citoesqueleto, movimiento celular, expresión genética y metabolismo. Existen diferentes rutas por las que el calcio interviene como:
Regulación de Proteínas G.
Regulación de los receptores de tirosincinasa.
Regulación de canales iónicos.
Existen dos caminos diferentes en los que el calcio puede regular proteínas:
Reconocimiento directo del calcio por la proteína
Unión del calcio al centro activo de una enzima.
Una de las interacciones mejor estudiadas del calcio con las proteínas es la regulación de la calmodulina por el calcio. La calmodulina por sí misma regula otras proteínas, o forma parte de grandes proteínas como por ejemplo la fosforilasa cinasa. El complejo calcio-calmodulina ejerce una función importante en la proliferación, mitosis y transducción de señal neuronal.
Óxido nítrico como segundo mensajero
El gas óxido nítrico (ON) es un radical libre que difunde a través de la membrana plasmática y afecta a las células vecinas. El ON se forma a partir de la arginina y el oxígeno por la enzima oxído nítrico sintetasa, con citrulina como sustrato. El ON funciona principalmente a través de receptores diana, la enzima soluble guanilato ciclasa, que cuando se activa produce el segundo mensajero guanosinmonofosfato cíclico (GMPc). El ON también puede actuar a través de la modificación covalente de proteínas o de su cofactor metálico. Algunas de estas modificaciones son reversibles y actúan a través de mecanismos de oxidación-reducción. En altas concentraciones el ON es tóxico, y se piensa que es el responsable de algunas lesiones después de un infarto. El ON realiza tres funciones principales:
Relajación de los vasos sanguíneos.
Regulación de la exocitosis de neurotransmisores.
Respuesta celular inmune.


{kind=link}